library(ggplot2)
library(grid)
te_ink <- "#16241d"; te_forest <- "#275139"; te_gold <- "#c9b458"
te_rust <- "#b5534e"; te_sage <- "#93a87f"
theme_te <- function() {
theme_minimal(base_size = 11) +
theme(panel.grid.minor = element_blank(),
panel.grid.major = element_line(colour = "grey90", linewidth = 0.3),
plot.title = element_text(colour = te_ink, face = "bold", size = 11),
plot.subtitle = element_text(colour = "grey35", size = 9),
axis.title = element_text(colour = te_ink, size = 9),
axis.text = element_text(colour = "grey30", size = 8),
legend.position = "bottom",
legend.title = element_blank(),
legend.text = element_text(size = 8))
}
two_panel <- function(p1, p2) {
grid.newpage()
pushViewport(viewport(layout = grid.layout(1, 2)))
print(p1, vp = viewport(layout.pos.row = 1, layout.pos.col = 1))
print(p2, vp = viewport(layout.pos.row = 1, layout.pos.col = 2))
}False positives in occupancy models
R
occupancy
imperfect detection
ecology tutorial
Why a five per cent misidentification rate wrecks an occupancy estimate, how the Royle-Link model repairs it, and the label swap that leaves two answers.
The standard occupancy model rests on an assumption so quiet that it is easy to miss: you can fail to see a species that is there, but you can never see a species that is not. Every correction the model applies runs in one direction. It assumes silence is evidence of nothing.
Break that assumption slightly, at a rate of one visit in twenty, and the model does not degrade gracefully. It reports occupancy nearly six times the truth, with a tight interval, and a goodness-of-fit test that raises no alarm.
A rare species and a fallible observer
Two hundred and fifty sites, five visits, a species present at one site in ten. When it is there you record it on 35% of visits. When it is not there you record it anyway on 5% of visits, because a similar call carries across the valley, or a footprint is ambiguous, or the automated classifier has a bad day.
set.seed(4274)
R <- 250L; K <- 5L
psi_t <- 0.10; p11_t <- 0.35; p10_t <- 0.05
z <- rbinom(R, 1L, psi_t)
Y <- matrix(rbinom(R * K, 1L, ifelse(rep(z, K) == 1, p11_t, p10_t)), R, K)
d <- rowSums(Y)
occupied <- sum(z)
detected <- sum(d > 0)
false_sites <- sum(d > 0 & z == 0)
naive <- mean(d > 0)The species truly occupies 22 of the 250 sites. It was recorded at least once at 73 sites, of which 55 hold no species at all. The naive estimate is 0.2920 against a truth of 0.10.
A careful analyst knows the naive estimate is biased and reaches for the correction.
ct <- list(maxit = 8000, reltol = 1e-12)
fit_nm <- function(nll, start, se = TRUE, ...) {
o <- optim(start, nll, ..., method = "Nelder-Mead", control = ct)
o <- optim(o$par, nll, ..., method = "Nelder-Mead", control = ct)
s <- rep(NA_real_, length(o$par))
if (se) {
H <- optimHess(o$par, nll, ...)
s <- tryCatch(sqrt(diag(solve(H))),
error = function(e) rep(NA_real_, length(o$par)))
}
list(par = o$par, se = s, nll = o$value,
aic = 2 * o$value + 2 * length(o$par))
}
occ_nll <- function(par, d, K) {
psi <- plogis(par[1]); p <- plogis(par[2])
pd <- psi * dbinom(0:K, K, p) + c(1 - psi, rep(0, K))
-sum(log(pmax(pd[d + 1], 1e-300)))
}
fp_nll <- function(par, d, K) {
psi <- plogis(par[1]); p11 <- plogis(par[2]); p10 <- plogis(par[3])
pd <- psi * dbinom(0:K, K, p11) + (1 - psi) * dbinom(0:K, K, p10)
-sum(log(pmax(pd[d + 1], 1e-300)))
}
f_occ <- fit_nm(occ_nll, c(0, 0), d = d, K = K)
f_fp <- fit_nm(fp_nll, c(0, qlogis(0.4), qlogis(0.05)), d = d, K = K)
psi_occ <- plogis(f_occ$par[1])
ci_occ <- plogis(f_occ$par[1] + c(-1.96, 1.96) * f_occ$se[1])
psi_fp <- plogis(f_fp$par[1])
ci_fp <- plogis(f_fp$par[1] + c(-1.96, 1.96) * f_fp$se[1])
implied_missed <- R * psi_occ - detectedThe standard occupancy model reports \(\hat\psi = 0.5752\), a 95% interval of [0.383, 0.747], against a truth of 0.10. It is not slightly wrong. It is wrong by a factor of 5.8.
The mechanism is worth stating plainly, because it is the opposite of what intuition suggests. The model sees 73 sites with detections and a fitted detection probability of only 0.132. Low detection means many occupied sites must have been missed entirely, so the model adds them: it infers roughly 71 occupied-but-never-detected sites, when the true number of such sites is 4. The correction for false negatives amplifies the false positives. A scatter of spurious single detections looks exactly like a real species that is hard to see, and the model responds by concluding the species is everywhere and shy.
The repair
Royle and Link (2006) add one parameter. Detection is a two-component mixture: occupied sites detect at \(p_{11}\), unoccupied sites detect at \(p_{10}\).
\[\Pr(d_i) = \psi \binom{K}{d_i} p_{11}^{d_i}(1-p_{11})^{K-d_i} + (1-\psi)\binom{K}{d_i} p_{10}^{d_i}(1-p_{10})^{K-d_i}\]
The standard model is the special case \(p_{10} = 0\). That is the entire difference.
p11_hat <- plogis(f_fp$par[2]); p10_hat <- plogis(f_fp$par[3])Fitted, it gives \(\hat\psi = 0.0925\) with an interval of [0.009, 0.540], \(\hat{p}_{11} = 0.305\) and \(\hat{p}_{10} = 0.053\), against truths of 0.35 and 0.05. The point estimate is now right and the interval is wide, which is the correct response to 250 sites and 22 of them occupied.
Notice what the AIC does here, though. The two models differ by 0.34. Between a model that says the species occupies 58% of sites and a model that says 9%, the fit statistics see almost nothing to choose. Both describe the detection frequencies about equally well, because both are two-component mixtures fitted to the same handful of numbers.
The label swap
The reason the AIC is helpless is exact, not approximate. Look at the likelihood again and swap the labels: replace \(\psi\) with \(1 - \psi\), and exchange \(p_{11}\) with \(p_{10}\). The two terms of the sum trade places, and the sum is unchanged.
\[\psi\, \text{Bin}(p_{11}) + (1-\psi)\,\text{Bin}(p_{10}) \;=\; (1-\psi)\,\text{Bin}(p_{10}) + \psi\, \text{Bin}(p_{11})\]
Every fit therefore has a mirror twin with identical likelihood, in which the species occupies 91% of sites, is detected there 5% of the time, and is falsely recorded 31% of the time at the sites where it is absent.
mirror <- c(-f_fp$par[1], f_fp$par[3], f_fp$par[2])
nll_mle <- fp_nll(f_fp$par, d, K)
nll_mirror <- fp_nll(mirror, d, K)
gap <- abs(nll_mle - nll_mirror)The negative log-likelihood at the mirror is 202.2843131186 against 202.2843131186 at the reported answer, a difference of 0.000e+00. The data are exactly, provably indifferent between the two.
Profile the likelihood over \(\psi\) and the invariance is visible as a shape: a double well, symmetric about one half, to machine precision.
prof_fp <- function(psi_fix, d, K, starts) {
f <- function(q) {
pd <- psi_fix * dbinom(0:K, K, plogis(q[1])) +
(1 - psi_fix) * dbinom(0:K, K, plogis(q[2]))
-sum(log(pmax(pd[d + 1], 1e-300)))
}
min(sapply(starts, function(s) {
o <- optim(s, f, method = "Nelder-Mead", control = ct)
optim(o$par, f, method = "Nelder-Mead", control = ct)$value
}))
}
psi_g <- seq(0.02, 0.98, length.out = 49)
lo_hi <- c(qlogis(0.4), qlogis(0.05)); hi_lo <- c(qlogis(0.05), qlogis(0.4))
prof <- sapply(psi_g, prof_fp, d = d, K = K, starts = list(lo_hi, hi_lo))
prof_one <- sapply(psi_g, prof_fp, d = d, K = K, starts = list(lo_hi))
dev_fp <- 2 * (prof - min(prof))
dev_one <- 2 * (prof_one - min(prof_one))
sym_gap <- max(abs(dev_fp - rev(dev_fp)))
sym_one <- max(abs(dev_one - rev(dev_one)))Profiled from two starts, one on each side of the swap, the deviance at \(\psi\) and at \(1-\psi\) differ by at most 2.274e-13 across the whole grid: exactly symmetric, as the algebra says it must be.
Profiled from a single start, they differ by up to 1.22. That gap is not a property of the likelihood, it is the optimiser falling into whichever of the two wells it started nearest, and reporting the result without complaint. A likelihood with a mirror twin needs a multi-start or it will lie to you quietly, in the same way that the Royle-Nichols likelihood does when BFGS walks it onto a boundary.
obs <- as.numeric(table(factor(d, levels = 0:K)))
e_occ <- R * (psi_occ * dbinom(0:K, K, plogis(f_occ$par[2])) +
c(1 - psi_occ, rep(0, K)))
e_fp <- R * (psi_fp * dbinom(0:K, K, p11_hat) + (1 - psi_fp) * dbinom(0:K, K, p10_hat))
dfa <- data.frame(d = rep(0:K, 2), count = c(e_occ, e_fp),
model = rep(c("standard occupancy", "false-positive model"),
each = K + 1))
pa <- ggplot() +
geom_col(data = data.frame(d = 0:K, count = obs), aes(d, count),
fill = te_sage, alpha = 0.55, width = 0.75) +
geom_line(data = dfa, aes(d, count, colour = model), linewidth = 0.7) +
geom_point(data = dfa, aes(d, count, colour = model), size = 1.6) +
scale_colour_manual(values = c("standard occupancy" = te_rust,
"false-positive model" = te_forest)) +
scale_x_continuous(breaks = 0:K) +
labs(title = "Both describe the data", x = "detections out of five", y = "sites",
subtitle = "bars: observed. the answers differ sixfold") +
theme_te()
pb <- ggplot(data.frame(psi = psi_g, dev = dev_fp), aes(psi, dev)) +
geom_vline(xintercept = 0.5, linetype = "dotted", colour = "grey45") +
geom_line(linewidth = 0.8, colour = te_forest) +
geom_vline(xintercept = psi_t, linetype = "dashed", colour = te_ink,
linewidth = 0.3) +
labs(title = "The likelihood has two answers", x = "occupancy",
y = "profile deviance",
subtitle = "dashed: truth. dotted: the axis of symmetry") +
theme_te()
two_panel(pa, pb)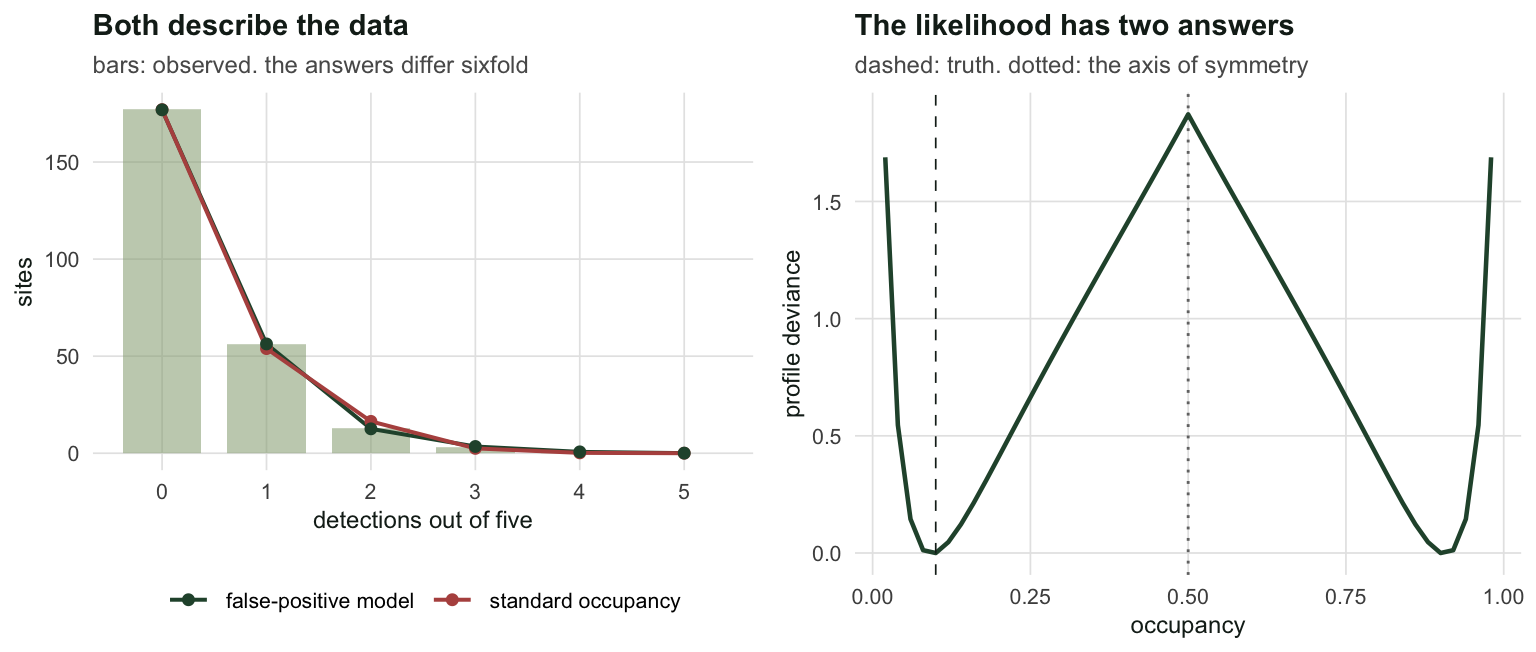
What picks the sensible one is not the likelihood but your prior conviction that a species is detected more reliably where it lives than where it does not, imposed as the constraint \(p_{11} > p_{10}\). That constraint is an assumption, and this model cannot test it. In a survey where the target is genuinely rarer than the confusable species, it is also the wrong assumption.
How bad, and when
One data set is an anecdote. Sweep the true occupancy across a range and refit, keeping everything else fixed.
psi_seq <- c(0.05, 0.10, 0.20, 0.30, 0.45, 0.60)
M <- 120L
sweep <- do.call(rbind, lapply(psi_seq, function(ps) {
est <- t(sapply(1:M, function(m) {
set.seed(70000L + m + round(ps * 1000))
zz <- rbinom(R, 1L, ps)
dd <- rowSums(matrix(rbinom(R * K, 1L,
ifelse(rep(zz, K) == 1, p11_t, p10_t)), R, K))
c(occ = plogis(fit_nm(occ_nll, c(0, 0), se = FALSE, d = dd, K = K)$par[1]),
fp = plogis(fit_nm(fp_nll, c(0, qlogis(0.4), qlogis(0.05)),
se = FALSE, d = dd, K = K)$par[1]))
}))
data.frame(psi = ps, model = c("standard occupancy", "false-positive model"),
bias = c(mean(est[, "occ"]) - ps, mean(est[, "fp"]) - ps))
}))
b_low <- sweep$bias[sweep$psi == 0.05 & sweep$model == "standard occupancy"]
b_high <- sweep$bias[sweep$psi == 0.60 & sweep$model == "standard occupancy"]lv <- c("naive", "standard occupancy", "false-positive model")
df1 <- data.frame(what = factor(lv, levels = lv), est = c(naive, psi_occ, psi_fp))
df_ci <- data.frame(what = factor(lv[-1], levels = lv),
lo = c(ci_occ[1], ci_fp[1]), hi = c(ci_occ[2], ci_fp[2]))
p1 <- ggplot(df1, aes(est, what)) +
geom_vline(xintercept = psi_t, linetype = "dashed", colour = te_ink,
linewidth = 0.4) +
geom_errorbarh(data = df_ci, aes(y = what, xmin = lo, xmax = hi),
inherit.aes = FALSE, height = 0.12, colour = te_forest,
linewidth = 0.7) +
geom_point(size = 2.6, colour = te_rust) +
labs(title = "One in twenty visits was wrong", x = "estimated occupancy", y = NULL,
subtitle = "dashed: the truth, 0.10") +
theme_te()Warning: `geom_errorbarh()` was deprecated in ggplot2 4.0.0.
ℹ Please use the `orientation` argument of `geom_errorbar()` instead.p2 <- ggplot(sweep, aes(psi, bias, colour = model)) +
geom_hline(yintercept = 0, linetype = "dashed", colour = te_ink, linewidth = 0.3) +
geom_line(linewidth = 0.8) +
geom_point(size = 1.7) +
scale_colour_manual(values = c("standard occupancy" = te_rust,
"false-positive model" = te_forest)) +
labs(title = "The rarer the species, the worse", x = "true occupancy",
y = "mean bias in estimated occupancy",
subtitle = "120 replicates per point") +
theme_te()
two_panel(p1, p2)`height` was translated to `width`.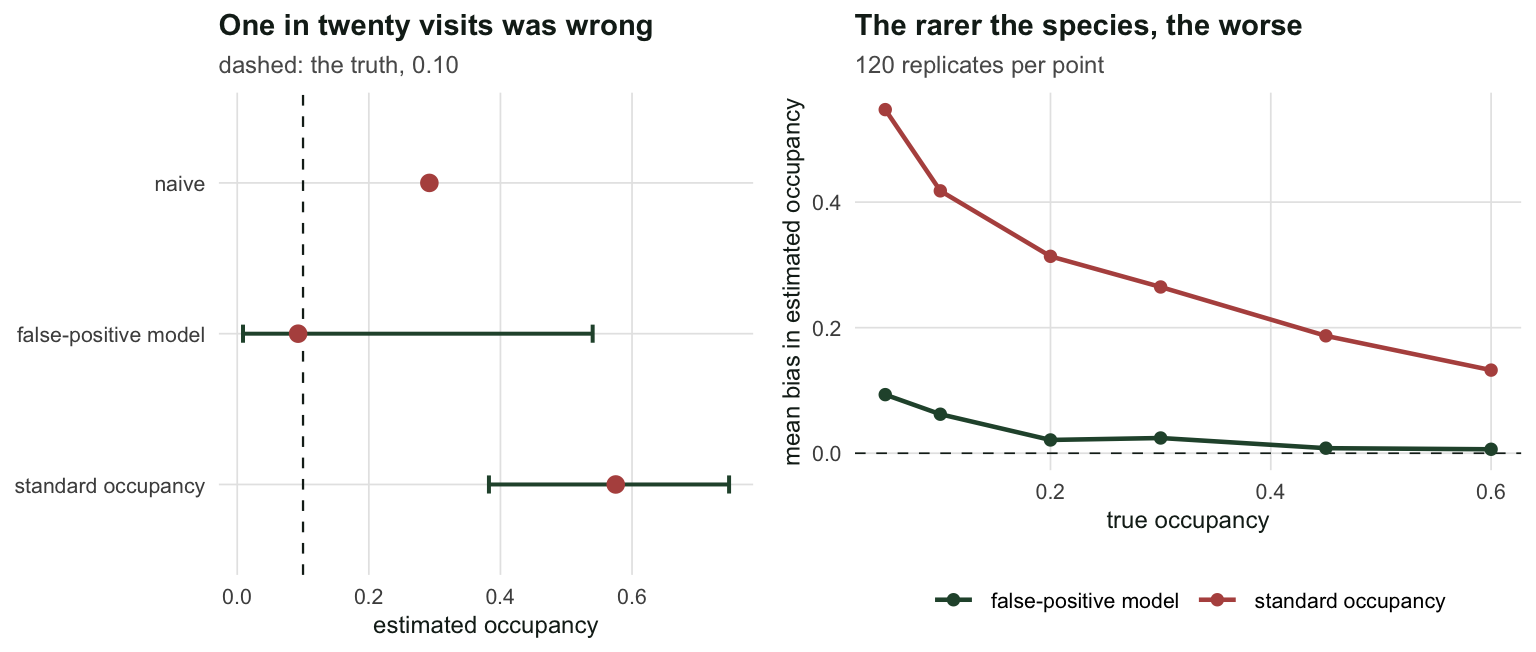
The bias runs from +0.547 at a true occupancy of 0.05 down to +0.132 at 0.60. This is the ugly part: the damage is worst exactly where occupancy models are most often used, which is on rare and cryptic species, and it shrinks to something ignorable for common species that nobody needed a model for. A 5% false-positive rate is not a small error at a site where the species occupies 5% of the landscape. It is comparable to the signal.
What actually fixes it
The label swap says the detection data alone cannot pin down which mixture component is the species. So bring something else.
Miller et al. (2011) formalise the practical answer: make a subset of detections certain. Record two classes of observation, one that can be wrong, such as an acoustic identification, and one that cannot, such as a specimen in hand or a photograph a second observer confirms. Certain detections cannot arise at unoccupied sites, so any site with one is occupied, full stop, and that anchors the mixture.
set.seed(4275)
b_t <- 0.40
W <- matrix(0L, R, K)
for (i in 1:R) for (k in 1:K) {
if (Y[i, k] == 1) {
W[i, k] <- if (z[i] == 1 && rbinom(1, 1, b_t) == 1) 2L else 1L
}
}
n1 <- rowSums(W == 1); n2 <- rowSums(W == 2); n0 <- K - n1 - n2
mil_nll <- function(par, n0, n1, n2, K) {
psi <- plogis(par[1]); p11 <- plogis(par[2])
p10 <- plogis(par[3]); b <- plogis(par[4])
occ <- (1 - p11)^n0 * (p11 * (1 - b))^n1 * (p11 * b)^n2
unocc <- ifelse(n2 > 0, 0, (1 - p10)^n0 * p10^n1)
-sum(log(pmax(psi * occ + (1 - psi) * unocc, 1e-300)))
}
f_mil <- fit_nm(mil_nll, c(0, qlogis(0.4), qlogis(0.05), 0),
n0 = n0, n1 = n1, n2 = n2, K = K)
psi_mil <- plogis(f_mil$par[1])
ci_mil <- plogis(f_mil$par[1] + c(-1.96, 1.96) * f_mil$se[1])
certain_sites <- sum(n2 > 0)With 40% of true detections upgraded to certain, 12 sites are now known to be occupied without any modelling at all. The three-state model returns \(\hat\psi = 0.1132\) with an interval of [0.048, 0.242], which is both correct and 2.7 times narrower than the two-parameter false-positive model managed on the same underlying sites.
That is the pattern worth taking away. The false-positive model does not extract more from the data; it declines to make a wrong assumption and pays for the honesty in width. What buys back the precision is not a better estimator but a field protocol that produces a class of observation which cannot lie. The decision that fixed this analysis was made before anyone opened R, and it is the same shape of decision as the one in the Royle-Nichols model, where counting something at a subset of sites is what turns an assumption into data.
References
Link W 2003 Biometrics 59(4):1123-1130 (doi:10.1111/j.0006-341X.2003.00129.x)
MacKenzie D, Nichols J, Lachman G, Droege S, Royle J, Langtimm C 2002 Ecology 83(8):2248-2255
Miller D, Nichols J, McClintock B, Campbell Grant E, Bailey L, Weir L 2011 Ecology 92(7):1422-1428 (doi:10.1890/10-1396.1)
Royle J, Link W 2006 Ecology 87(4):835-841