---
title: "Mark resight abundance estimation in R"
description: "Poisson log-normal mark resight from scratch: why individual heterogeneity leaves the estimate alone but destroys the interval, and what the marked animals cannot tell you."
date: "2026-07-17 15:00"
categories: [R, abundance, capture-recapture, imperfect detection, ecology tutorial]
image: thumbnail.png
image-alt: "Two line panels showing that the naive coefficient of variation stays flat while the true one rises with heterogeneity, collapsing interval coverage"
---
Mark resight sits between two methods you already know. Like [closed capture-recapture](../closed-capture-recapture/), it uses marked animals to estimate how many you missed. Unlike it, the marked total is known exactly, because you put the collars on yourself, and the resightings are counts rather than captures: you see an animal, you note whether it is marked, and you move on. An individual can be seen five times in one survey.
That last difference changes the statistics more than it looks, and in a direction that surprises people who arrive from capture-recapture. The received wisdom there is that individual heterogeneity in detection biases abundance downwards. Here it does not bias the estimate at all. It destroys the confidence interval instead, and it does so silently.
## The data
Four hundred animals, eighty collared, twelve resighting occasions. Sighting rates vary between individuals: some animals live near the road, some do not.
```{r data}
set.seed(3140)
sim_mr <- function(N = 400, n = 80, J = 12, lam = 0.35, sigma = 0.8, seed = 1) {
set.seed(seed)
# individual sighting rate, log-normal, with mean fixed at lam
alpha <- rlnorm(N, log(lam) - sigma^2 / 2, sigma)
mk <- sample(N, n)
y <- matrix(rpois(N * J, rep(alpha, J)), N, J)
list(N = N, n = n, J = J,
ym = y[mk, , drop = FALSE], # marked: identified individually
Tu = sum(y[-mk, , drop = FALSE])) # unmarked: a count, nothing more
}
d <- sim_mr(seed = 7)
ytot <- rowSums(d$ym)
p314_Tm <- sum(ytot); p314_Tu <- d$Tu
c(true_N = d$N, marked = d$n, occasions = d$J,
marked_sightings = p314_Tm, unmarked_sightings = p314_Tu)
```
Note what the unmarked animals give you: a single number per occasion. You cannot tell whether `r p314_Tu` unmarked sightings came from `r p314_Tu` different animals or from a handful of very visible ones. Everything the analysis learns about that distribution, it learns from the collared animals.
## The estimator
The logic is a rate argument, not a capture argument. Marked animals were seen `r p314_Tm` times over `r d$n` animals and `r d$J` occasions, so the sighting rate per animal per occasion is that ratio. Unmarked animals were seen `r p314_Tu` times at the same rate, so divide.
```{r estimator}
lam_hat <- sum(ytot) / (d$n * d$J)
U_hat <- d$Tu / (d$J * lam_hat)
N_hat <- d$n + U_hat
p314_lam <- lam_hat; p314_N <- N_hat
c(rate = lam_hat, unmarked_est = U_hat, N_hat = N_hat, true = d$N)
```
This is the moment estimator, and it is algebraically the mark ratio: `N = n * (total sightings) / (marked sightings)`. Now the part that matters.
The expected total number of sightings is the number of animals times the number of occasions times the *mean* sighting rate. Heterogeneity spreads the individual rates around that mean, but it does not move the mean, and the mean is all the estimator uses. This is where mark resight parts company with capture-recapture. There, the quantity that matters is the probability of being caught *at least once*, and heterogeneity pushes that down: the shy animals form a class that is never seen, and the estimate misses them. Here, a shy animal contributes few sightings and a bold animal contributes many, and the arithmetic does not care which is which as long as the total is right.
So the point estimate survives. The interval does not.
## Where the variance hides
Treat the sightings as Poisson counts and the standard errors are easy. They are also wrong, and the marked animals will tell you so if you ask.
```{r overdispersion}
p314_mean <- mean(ytot); p314_var <- var(ytot)
p314_ratio <- p314_var / p314_mean
c(mean_of_individual_totals = p314_mean,
variance_of_individual_totals = p314_var,
ratio = p314_ratio)
```
Under a pure Poisson process with a common rate, an individual's total over twelve occasions is Poisson, and its variance equals its mean. The observed ratio is `r sprintf("%.2f", p314_ratio)`. That excess is heterogeneity, measured directly, and it inflates the variance of every count in the study, including the unmarked total that the estimator divides.
## Poisson log-normal, from scratch
To use that excess we need a model for it. The standard choice is a log-normal individual effect: each animal has its own rate, the log of which is normal. The likelihood for one animal's total needs the latent rate integrated out, which is a one-dimensional Gaussian integral, and Gauss-Hermite quadrature does it in a few nodes.
```{r gh}
gh_nodes <- function(K) { # Golub-Welsch for the weight exp(-x^2)
i <- 1:(K - 1)
Jm <- matrix(0, K, K)
Jm[cbind(i, i + 1)] <- sqrt(i / 2)
Jm[cbind(i + 1, i)] <- sqrt(i / 2)
e <- eigen(Jm, symmetric = TRUE)
o <- order(e$values)
list(x = e$values[o], w = sqrt(pi) * e$vectors[1, o]^2)
}
GH <- gh_nodes(20)
# validate on integrals we know: E[Z^2] = 1 and E[exp(Z)] = exp(1/2)
p314_ez2 <- sum(GH$w / sqrt(pi) * (sqrt(2) * GH$x)^2)
p314_eez <- sum(GH$w / sqrt(pi) * exp(sqrt(2) * GH$x))
c(E_Z2 = p314_ez2, E_expZ = p314_eez, target = exp(0.5))
```
Both match to machine precision, so the quadrature is doing what it claims. That check is not ceremony: a quadrature rule with the nodes built wrong returns a plausible number rather than an error, and it will hand you a rate estimate that is quietly off by twenty percent.
```{r pln}
mk_tab <- function(y) { tb <- table(y); list(v = as.integer(names(tb)), k = as.integer(tb)) }
nll_pln <- function(par, tab, J) {
lam_i <- exp(par[1] + sqrt(2) * exp(par[2]) * GH$x) # rate at each node
M <- outer(tab$v, J * lam_i, dpois) # unique totals x nodes
-sum(tab$k * log(as.numeric(M %*% (GH$w / sqrt(pi)))))
}
tab <- mk_tab(ytot)
f <- optim(c(log(0.3), log(0.8)), nll_pln, tab = tab, J = d$J, method = "BFGS")
mu_h <- f$par[1]; sg_h <- exp(f$par[2])
lam_pln <- exp(mu_h + sg_h^2 / 2)
p314_sg <- sg_h; p314_lam_pln <- lam_pln
c(sigma = sg_h, mean_rate = lam_pln, moment_rate = lam_hat)
```
Sigma comes back at `r sprintf("%.3f", p314_sg)` against a true 0.8. The mean rate agrees with the moment estimator to the third decimal, which is the point: the model is not there to improve the rate. It is there to measure the spread around it, so the variance can be computed honestly.
```{r variance}
J <- d$J
cv2 <- exp(sg_h^2) - 1 # squared CV of the individual rate
var_yi <- J * lam_hat + J^2 * lam_hat^2 * cv2 # variance of one animal's total
v_lam <- var_yi / (d$n * J^2)
v_Tu_naive <- d$Tu # Poisson: variance equals the count
v_Tu_hon <- U_hat * (J * lam_hat + J^2 * lam_hat^2 * cv2)
se_naive <- sqrt(v_Tu_naive / (J * lam_hat)^2 + (U_hat / lam_hat)^2 * (lam_hat / (d$n * J)))
se_hon <- sqrt(v_Tu_hon / (J * lam_hat)^2 + (U_hat / lam_hat)^2 * v_lam)
p314_se_n <- se_naive; p314_se_h <- se_hon
c(se_naive = se_naive, se_honest = se_hon, ratio = se_hon / se_naive)
```
## Three hundred surveys, counted
One dataset proves nothing about coverage. Run the study repeatedly against a known truth.
```{r mc}
fit_one <- function(d) {
yt <- rowSums(d$ym); J <- d$J; n <- d$n
lam <- sum(yt) / (n * J)
U <- d$Tu / (J * lam); N <- n + U
se_n <- sqrt(d$Tu / (J * lam)^2 + (U / lam)^2 * (lam / (n * J)))
tb <- mk_tab(yt)
ff <- optim(c(log(lam), log(0.8)), nll_pln, tab = tb, J = J, method = "BFGS")
sg <- exp(ff$par[2]); cv2 <- exp(sg^2) - 1
v_lam <- (J * lam + J^2 * lam^2 * cv2) / (n * J^2)
v_Tu <- U * (J * lam + J^2 * lam^2 * cv2)
se_h <- sqrt(v_Tu / (J * lam)^2 + (U / lam)^2 * v_lam)
c(N = N, se_naive = se_n, se_hon = se_h, sigma = sg)
}
M <- 300
res <- t(sapply(1:M, function(i) fit_one(sim_mr(seed = 8000 + i))))
p314_bias <- 100 * (mean(res[, "N"]) / 400 - 1)
p314_truecv <- sd(res[, "N"]) / mean(res[, "N"])
p314_cv_n <- mean(res[, "se_naive"] / res[, "N"])
p314_cv_h <- mean(res[, "se_hon"] / res[, "N"])
p314_sg_mc <- mean(res[, "sigma"])
p314_cov_n <- mean(abs(res[, "N"] - 400) <= 1.96 * res[, "se_naive"])
p314_cov_h <- mean(abs(res[, "N"] - 400) <= 1.96 * res[, "se_hon"])
data.frame(replicates = M, rel_bias_pct = p314_bias, mean_sigma = p314_sg_mc,
cv_actual = p314_truecv, cv_naive = p314_cv_n, cv_honest = p314_cv_h,
cover_naive = p314_cov_n, cover_honest = p314_cov_h)
```
Relative bias `r sprintf("%+.2f%%", p314_bias)`. The estimator is essentially unbiased, as promised.
Across replicates it varies with a coefficient of variation of `r sprintf("%.3f", p314_truecv)`. The naive formula reports `r sprintf("%.3f", p314_cv_n)`, less than half of it, and a nominal 95% interval covers the truth `r sprintf("%.1f%%", 100 * p314_cov_n)` of the time. The overdispersion-corrected formula reports `r sprintf("%.3f", p314_cv_h)`, essentially the truth, and covers `r sprintf("%.1f%%", 100 * p314_cov_h)`.
## The fingerprint
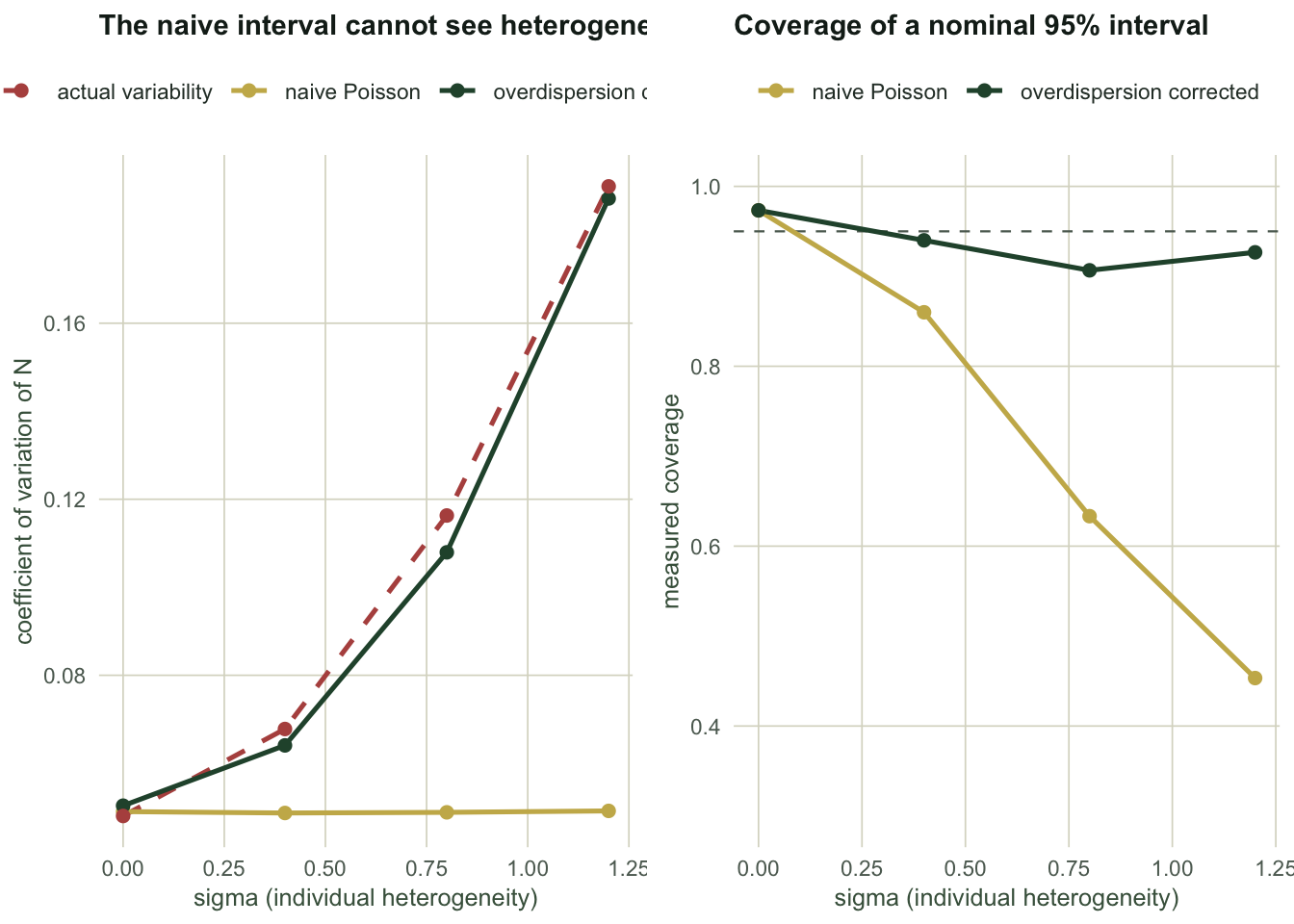
Now sweep the heterogeneity from none to severe. This is the figure that makes the argument.
```{r sweep}
sweep_sigma <- function(sg, R = 150) {
r <- t(sapply(1:R, function(i) fit_one(sim_mr(sigma = sg, seed = 9000 + i))))
data.frame(sigma = sg,
bias = 100 * (mean(r[, "N"]) / 400 - 1),
cv_true = sd(r[, "N"]) / mean(r[, "N"]),
cv_naive = mean(r[, "se_naive"] / r[, "N"]),
cv_honest = mean(r[, "se_hon"] / r[, "N"]),
cover_naive = mean(abs(r[, "N"] - 400) <= 1.96 * r[, "se_naive"]),
cover_honest = mean(abs(r[, "N"] - 400) <= 1.96 * r[, "se_hon"]))
}
sw <- do.call(rbind, lapply(c(0, 0.4, 0.8, 1.2), sweep_sigma))
p314_sw <- sw
print(round(sw, 4), row.names = FALSE)
```
Read the `cv_naive` column downwards. It is `r sprintf("%.4f", sw$cv_naive[1])`, then `r sprintf("%.4f", sw$cv_naive[2])`, then `r sprintf("%.4f", sw$cv_naive[3])`, then `r sprintf("%.4f", sw$cv_naive[4])`. It does not move. Now read `cv_true` beside it: `r sprintf("%.4f", sw$cv_true[1])` rising to `r sprintf("%.4f", sw$cv_true[4])`, roughly a factor of `r sprintf("%.1f", sw$cv_true[4] / sw$cv_true[1])`.
The naive interval has no channel through which heterogeneity could reach it. It is computed from counts, and the counts look the same whether they came from four hundred identical animals or from four hundred wildly different ones. Coverage falls from `r sprintf("%.3f", sw$cover_naive[1])` to `r sprintf("%.3f", sw$cover_naive[4])` while the formula reports the same precision throughout. At the far end you are quoting a 95% interval that is really a 43% interval.
The corrected version tracks: `cv_honest` climbs from `r sprintf("%.4f", sw$cv_honest[1])` to `r sprintf("%.4f", sw$cv_honest[4])`, following the truth, and coverage stays near nominal.
```{r fig-sweep}
#| echo: false
#| fig-cap: "Left: the naive coefficient of variation is flat across heterogeneity while the true one rises. Right: the resulting coverage of a nominal 95% interval."
#| fig-alt: "Two panels against sigma from zero to 1.2. Left panel: a flat gold line near 0.05 for the naive coefficient of variation, a rising green line for the honest one, and a rising dashed line for the true variability, with the gold line diverging further at each step. Right panel: coverage, the gold naive line falling from 0.97 to about 0.43 while the green honest line stays between 0.88 and 0.98 near the dashed nominal 0.95."
library(ggplot2)
theme_te <- function() {
theme_minimal(base_size = 11) +
theme(panel.grid.minor = element_blank(),
panel.grid.major = element_line(colour = "#dad9ca", linewidth = 0.3),
plot.title = element_text(colour = "#16241d", face = "bold", size = 11),
axis.title = element_text(colour = "#46604a", size = 9.5),
axis.text = element_text(colour = "#5d6b61", size = 8.5),
legend.position = "top", legend.title = element_blank(),
legend.text = element_text(colour = "#2c3a31", size = 8.5))
}
d1 <- rbind(
data.frame(s = sw$sigma, v = sw$cv_naive, k = "naive Poisson"),
data.frame(s = sw$sigma, v = sw$cv_honest, k = "overdispersion corrected"),
data.frame(s = sw$sigma, v = sw$cv_true, k = "actual variability"))
pA <- ggplot(d1, aes(s, v, colour = k, linetype = k)) +
geom_line(linewidth = 0.9) + geom_point(size = 2) +
scale_colour_manual(values = c("naive Poisson" = "#c9b458",
"overdispersion corrected" = "#275139",
"actual variability" = "#b5534e")) +
scale_linetype_manual(values = c("naive Poisson" = "solid",
"overdispersion corrected" = "solid",
"actual variability" = "dashed")) +
labs(title = "The naive interval cannot see heterogeneity",
x = "sigma (individual heterogeneity)", y = "coefficient of variation of N") +
theme_te()
d2 <- rbind(data.frame(s = sw$sigma, v = sw$cover_naive, k = "naive Poisson"),
data.frame(s = sw$sigma, v = sw$cover_honest, k = "overdispersion corrected"))
pB <- ggplot(d2, aes(s, v, colour = k)) +
geom_hline(yintercept = 0.95, linetype = "dashed", colour = "#5d6b61", linewidth = 0.4) +
geom_line(linewidth = 0.9) + geom_point(size = 2) +
scale_colour_manual(values = c("naive Poisson" = "#c9b458",
"overdispersion corrected" = "#275139")) +
scale_y_continuous(limits = c(0.3, 1)) +
labs(title = "Coverage of a nominal 95% interval",
x = "sigma (individual heterogeneity)", y = "measured coverage") +
theme_te()
grid::grid.newpage()
grid::pushViewport(grid::viewport(layout = grid::grid.layout(1, 2)))
print(pA, vp = grid::viewport(layout.pos.row = 1, layout.pos.col = 1))
print(pB, vp = grid::viewport(layout.pos.row = 1, layout.pos.col = 2))
```
## The honest limit
Everything above rests on one assumption that no diagnostic in the analysis can test: that the marked animals are a fair sample of the sighting process for the unmarked ones.
They are the only animals you can measure individually. Sigma comes from them; the overdispersion correction comes from them; the mean rate that divides the unmarked total comes from them. If collaring changes how visible an animal is, in either direction, all of it is measured on a population that does not exist outside the collars.
Here is what that costs. Suppose the collar makes an animal slightly less likely to be noticed, with a multiplier on its sighting rate.
```{r collar}
sim_eta <- function(N = 400, n = 80, J = 12, lam = 0.35, sigma = 0.8, eta = 1, seed = 1) {
set.seed(seed)
alpha <- rlnorm(N, log(lam) - sigma^2 / 2, sigma)
mk <- sample(N, n)
a2 <- alpha; a2[mk] <- a2[mk] * eta # the collar effect, marked only
y <- matrix(rpois(N * J, rep(a2, J)), N, J)
list(N = N, n = n, J = J, ym = y[mk, , drop = FALSE], Tu = sum(y[-mk, , drop = FALSE]))
}
collar_row <- function(eta, R = 120) {
r <- t(sapply(1:R, function(i) {
dd <- sim_eta(eta = eta, seed = 7000 + i)
out <- fit_one(dd)
c(out, ratio = var(rowSums(dd$ym)) / mean(rowSums(dd$ym)))
}))
data.frame(eta = eta, mean_N = mean(r[, "N"]), bias = 100 * (mean(r[, "N"]) / 400 - 1),
var_mean_ratio = mean(r[, "ratio"]), sigma_hat = mean(r[, "sigma"]))
}
cs <- do.call(rbind, lapply(c(1.0, 0.9, 0.8, 0.7), collar_row))
p314_cs <- cs
print(round(cs, 3), row.names = FALSE)
```
A ten percent reduction in sightability produces a `r sprintf("%+.0f%%", cs$bias[2])` error in abundance. Thirty percent produces `r sprintf("%+.0f%%", cs$bias[4])`. The direction is fixed: undercounting the marked animals makes the estimated rate too low, which makes the unmarked population look larger.
And now the part to remember. Look at `sigma_hat` down that table: `r sprintf("%.3f", cs$sigma_hat[1])`, `r sprintf("%.3f", cs$sigma_hat[2])`, `r sprintf("%.3f", cs$sigma_hat[3])`, `r sprintf("%.3f", cs$sigma_hat[4])`. It does not move. The model fits, the heterogeneity is recovered correctly, the overdispersion ratio stays in a sensible range, the interval has honest coverage around the wrong number. Every diagnostic in this post passes on data that is off by a third.
This is not a flaw in the estimator. It is the shape of the information: a rate measured on one group, applied to another, with nothing in the data connecting the two. The defence is design, not analysis. Collars that do not change behaviour, a marking period long enough for animals to settle, and a marked sample drawn the same way as the population you are counting.
## What to report
The abundance estimate, the sigma, the variance-to-mean ratio of the marked totals, and the interval computed with the overdispersion in it. If sigma is near zero the correction costs you nothing, as the sweep shows at the top row. If it is not, the correction is the difference between an interval and a decoration.
## Where to go next
If your animals are individually identifiable at every encounter, you want [closed capture-recapture](../closed-capture-recapture/), where the heterogeneity problem bites the estimate rather than the interval. If none of them are marked, [the random encounter model](../random-encounter-model-camera-traps/) gets density from sighting rates alone, at the cost of needing animal speed. If you can use the locations of the sightings as well as their counts, [spatial capture-recapture](../spatial-capture-recapture-basics/) uses far more of the information in the same field effort.
## References
McClintock, White, Antolin & Tripp 2009 Biometrics 65(1):237-246 (10.1111/j.1541-0420.2008.01047.x)
McClintock, White & Burnham 2006 Journal of Agricultural, Biological, and Environmental Statistics 11(3):231-248 (10.1198/108571106X129171)
McClintock & White 2009 Ecology 90(2):313-320 (10.1890/08-0973.1)
McClintock, White & Pryde 2019 Biometrics 75(3):799-809 (10.1111/biom.13058)
Bowden & Kufeld 1995 Journal of Wildlife Management 59:840-851
Royle, Chandler, Sollmann & Gardner 2014 Spatial Capture-Recapture, ISBN 978-0-12-405939-9
## Related tutorials
- [Closed population capture-recapture in R](../closed-capture-recapture/)
- [The random encounter model for camera traps](../random-encounter-model-camera-traps/)
- [Spatial capture-recapture from scratch](../spatial-capture-recapture-basics/)
- [N-mixture models for abundance](../n-mixture-models-abundance/)