library(ggplot2)
library(grid)
res_raw <- function(x, y) {
1.5 * exp(-((x - 36)^2 + (y - 64)^2) / (2 * 22^2)) +
1.2 * exp(-((x - 70)^2 + (y - 38)^2) / (2 * 24^2)) +
(-0.00034) * ((x - 50)^2 + (y - 50)^2)
}
gx <- seq(0, 100, length.out = 120)
grid_df <- expand.grid(x = gx, y = gx)
grid_df$raw <- res_raw(grid_df$x, grid_df$y)
mu_r <- mean(grid_df$raw); sd_r <- sd(grid_df$raw)
resource <- function(x, y) (res_raw(x, y) - mu_r) / sd_r
rvm <- function(n, mu, kappa) {
out <- numeric(n); a <- 1 + sqrt(1 + 4 * kappa^2)
b <- (a - sqrt(2 * a)) / (2 * kappa); r <- (1 + b^2) / (2 * b)
for (i in seq_len(n)) {
repeat {
u1 <- runif(1); u2 <- runif(1); u3 <- runif(1)
z <- cos(pi * u1); f <- (1 + r * z) / (r + z); cc <- kappa * (r - f)
if (cc * (2 - cc) - u2 > 0 || log(cc / u2) + 1 - cc >= 0) break
}
out[i] <- sign(u3 - 0.5) * acos(f) + mu
}
((out + pi) %% (2 * pi)) - pi
}Availability sampling for step selection
movement ecology
habitat selection
R
ecology tutorial
How many available steps per used step, and from what distribution: more steps buy precision, while iSSA barely depends on the tentative kernel.
Every step selection analysis rests on a choice that is easy to make without thinking: how many available steps to pair with each used step, and what distribution to draw them from. This is the movement version of the question that haunts presence-background models, how many background points and from where. The two halves have different answers. The number of available steps controls how precisely you pin down the coefficients. The distribution they come from controls whether the coefficients are biased at all, and that is where the integrated approach earns its keep.
A track to sample against
We reuse the generative model from the earlier posts: an animal with a known movement kernel selecting for a resource with known strength. One track, so that everything downstream is a property of the sampling, not of a fresh simulation each time.
k_true <- 2.0; th_true <- 6.0; kap_true <- 0.9; beta_true <- 0.9
n <- 1500; M <- 300
set.seed(3141)
pos <- matrix(NA_real_, n + 1, 2); pos[1, ] <- c(46, 52)
heading <- numeric(n + 1); heading[1] <- runif(1, -pi, pi)
for (i in 1:n) {
L <- rgamma(M, shape = k_true, scale = th_true); a <- rvm(M, 0, kap_true)
ang <- heading[i] + a
ex <- pos[i, 1] + L * cos(ang); ey <- pos[i, 2] + L * sin(ang)
w <- exp(beta_true * resource(ex, ey)); j <- sample.int(M, 1, prob = w)
pos[i + 1, ] <- c(ex[j], ey[j]); heading[i + 1] <- ang[j]
}
dx <- diff(pos[, 1]); dy <- diff(pos[, 2])
obs_l <- sqrt(dx^2 + dy^2); bearing <- atan2(dy, dx)
obs_turn <- ((diff(bearing) + pi) %% (2 * pi)) - pi
use_idx <- 2:n; n_str <- length(use_idx)
m1 <- mean(obs_l); v1 <- var(obs_l); k0 <- m1^2 / v1; th0 <- v1 / m1; lam0 <- 1 / th0
Cc <- mean(cos(obs_turn)); Ss <- mean(sin(obs_turn)); Rbar <- sqrt(Cc^2 + Ss^2); mu0 <- atan2(Ss, Cc)
kap0 <- if (Rbar < 0.53) 2 * Rbar + Rbar^3 + 5 * Rbar^5 / 6 else
if (Rbar < 0.85) -0.4 + 1.39 * Rbar + 0.43 / (1 - Rbar) else
1 / (Rbar^3 - 4 * Rbar^2 + 3 * Rbar)How many available steps
A basic step selection function fits habitat as the only covariate, one used step against K available steps drawn from the tentative kernel. To separate the sampling noise from everything else, we fit the same track many times, redrawing the available steps each time, and watch how the estimate behaves as K grows.
fit_ssf <- function(Kk, sc, sh = k0, kp = kap0, seed) {
set.seed(seed)
Zc <- resource(pos[use_idx + 1, 1], pos[use_idx + 1, 2])
Zm <- matrix(NA_real_, n_str, Kk)
for (s in seq_along(use_idx)) {
i <- use_idx[s]
Ls <- rgamma(Kk, shape = sh, scale = sc); As <- rvm(Kk, mu0, kp)
ang <- heading[i] + As
Zm[s, ] <- resource(pos[i, 1] + Ls * cos(ang), pos[i, 2] + Ls * sin(ang))
}
Z <- cbind(Zc, Zm)
o <- optim(0, function(b) { e <- Z * b; -sum(e[, 1] - log(rowSums(exp(e)))) },
method = "BFGS", hessian = TRUE, control = list(reltol = 1e-9))
c(est = o$par, se = sqrt(1 / o$hessian[1, 1]))
}
Kset <- c(1, 2, 5, 10, 20, 50, 100); R <- 40
setA <- data.frame()
for (Kk in Kset) {
ests <- ses <- numeric(R)
for (rr in 1:R) {
fr <- fit_ssf(Kk, sc = th0, seed = 1000 * Kk + rr)
ests[rr] <- fr["est"]; ses[rr] <- fr["se"]
}
setA <- rbind(setA, data.frame(K = Kk, mean_est = mean(ests),
sd_est = sd(ests), mean_se = mean(ses)))
}Two things move as K rises. The spread of the estimate across redraws, which is pure sampling noise from the availability set, falls from 0.053 at a single available step to 0.006 at a hundred. The model’s own standard error falls too, but it does not fall forever: it starts near 0.078 and settles around 0.053. That floor is set by the number of used steps, not by K. Once you have drawn enough available steps to integrate the availability well, adding more cannot tell you anything the 1499 observed steps do not already contain.
ggplot(setA, aes(K, mean_est)) +
geom_hline(yintercept = beta_true, linetype = "dashed", colour = "#b5534e") +
geom_ribbon(aes(ymin = mean_est - sd_est, ymax = mean_est + sd_est),
fill = "#93a87f", alpha = 0.35) +
geom_line(colour = "#275139", linewidth = 0.6) +
geom_point(colour = "#275139", size = 2) +
scale_x_log10(breaks = Kset) +
annotate("text", x = 1.15, y = beta_true + 0.02, label = "true beta",
colour = "#b5534e", size = 3, hjust = 0) +
labs(x = "available steps per used step (K, log scale)",
y = "habitat coefficient", title = "Precision grows with K, then plateaus") +
theme_minimal(base_size = 11) +
theme(panel.grid.minor = element_blank(),
plot.title = element_text(size = 10, colour = "#46604a"))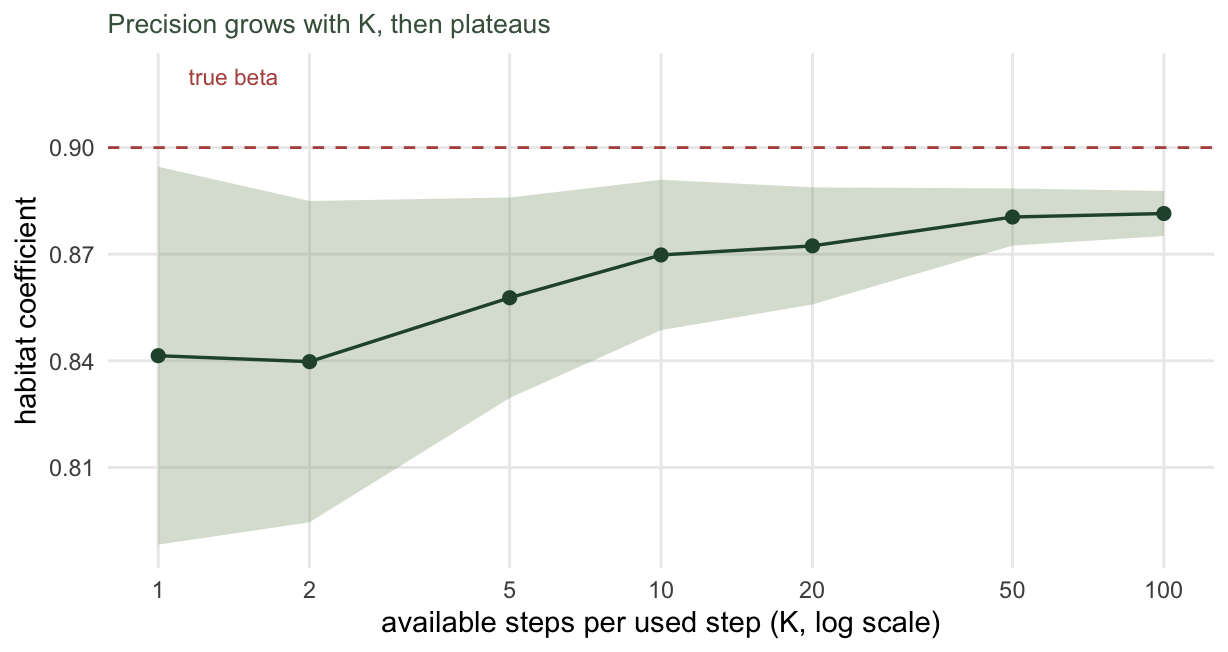
Notice where the curve settles: a little under the true value of 0.9. More available steps make the estimate tighter and steadier, but they do not move it onto the truth. The residual gap is bias from the contaminated tentative kernel, and no amount of sampling fixes bias. That is the second question.
What distribution to draw from
The available steps here come from a gamma fitted to the observed steps, which the earlier post showed is already shaped by selection. Does the choice matter? We take three tentative step-length distributions, one too short, one matched to the observed steps, one too long, and fit each with a basic step selection function and with the integrated version that carries movement covariates.
fit_issa <- function(sc, Kk = 15, seed = 77) {
set.seed(seed)
p <- 4; X <- matrix(NA_real_, n_str * (Kk + 1), p); row <- 1
for (s in seq_along(use_idx)) {
i <- use_idx[s]
lu <- obs_l[i]; tu <- obs_turn[i - 1]
X[row, ] <- c(resource(pos[i + 1, 1], pos[i + 1, 2]), log(lu), lu, cos(tu)); row <- row + 1
Ls <- rgamma(Kk, shape = k0, scale = sc); As <- rvm(Kk, mu0, kap0)
ang <- heading[i] + As
ex <- pos[i, 1] + Ls * cos(ang); ey <- pos[i, 2] + Ls * sin(ang)
X[row:(row + Kk - 1), ] <- cbind(resource(ex, ey), log(Ls), Ls, cos(As)); row <- row + Kk
}
nll <- function(b) { e <- as.vector(X %*% b); em <- matrix(e, nrow = Kk + 1)
-sum(em[1, ] - log(colSums(exp(em)))) }
optim(rep(0, p), nll, method = "BFGS", control = list(reltol = 1e-9))$par[1]
}
scales <- c(0.6, 1.0, 1.6) * th0; labs <- c("too short", "matched", "too long")
setB <- data.frame()
for (m in seq_along(scales)) {
b_basic <- fit_ssf(15, sc = scales[m], seed = 5000 + m)["est"]
b_issa <- fit_issa(scales[m], Kk = 15, seed = 6000 + m)
setB <- rbind(setB, data.frame(tentative = labs[m], basic = b_basic, issa = b_issa))
}The basic model is at the mercy of the choice. Its habitat coefficient walks from 0.81 under the too-short kernel to 0.89 under the too-long one, and all three sit below the true 0.9. The integrated model barely moves: 0.91, 0.95 and 0.95, all hugging the truth. The movement covariates absorb whatever the tentative distribution gets wrong, so the habitat coefficient stops depending on it.
setB_long <- rbind(
data.frame(tentative = setB$tentative, method = "basic SSF", est = setB$basic),
data.frame(tentative = setB$tentative, method = "iSSA", est = setB$issa))
setB_long$tentative <- factor(setB_long$tentative, levels = labs)
ggplot(setB_long, aes(tentative, est, colour = method, group = method)) +
geom_hline(yintercept = beta_true, linetype = "dashed", colour = "#b5534e") +
geom_line(linewidth = 0.6) + geom_point(size = 2.8) +
scale_colour_manual(values = c("basic SSF" = "#b5534e", "iSSA" = "#275139")) +
annotate("text", x = 0.7, y = beta_true + 0.03, label = "true beta",
colour = "#b5534e", size = 3, hjust = 0) +
labs(x = "tentative step-length distribution", y = "habitat coefficient",
colour = NULL, title = "iSSA holds steady across tentatives; basic SSF does not") +
theme_minimal(base_size = 11) +
theme(panel.grid.minor = element_blank(), legend.position = c(0.5, 0.14),
legend.direction = "horizontal",
legend.background = element_rect(fill = "#ffffff", colour = NA),
plot.title = element_text(size = 10, colour = "#46604a"))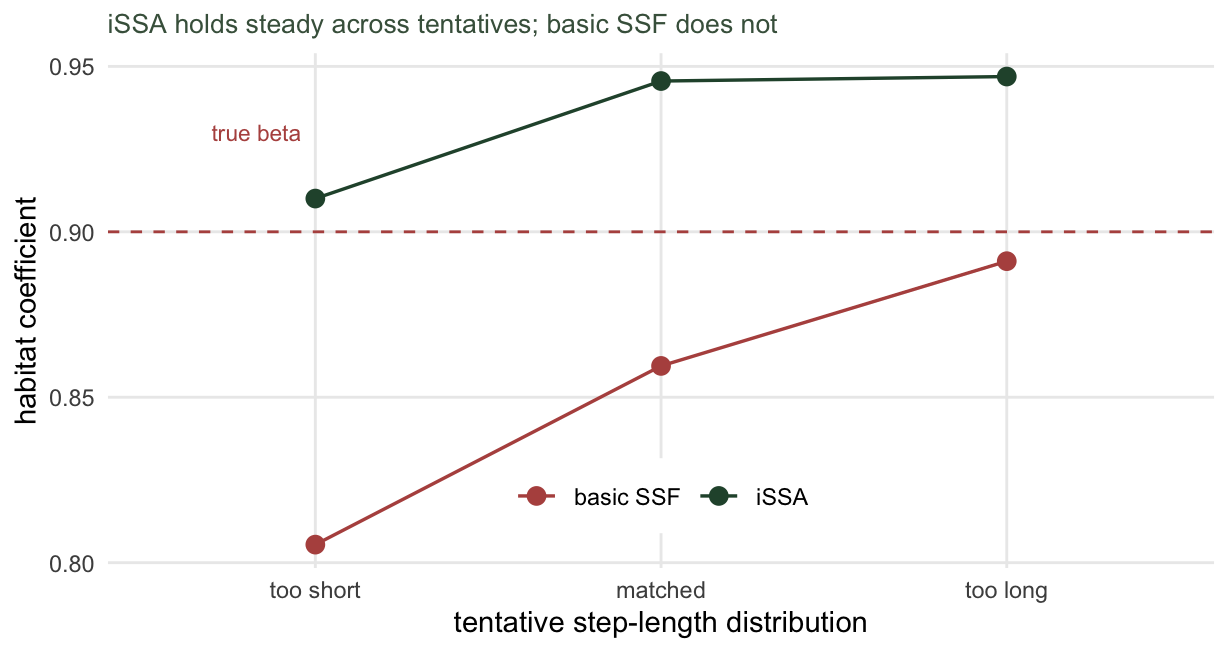
This steadiness has limits. The integrated model corrects the parametric form of the tentative kernel, so it needs available steps that actually cover the range the animal walked. A tentative so narrow that it never proposes a long step leaves the correction extrapolating into empty space, and eventually even the integrated estimate suffers. The three kernels here all overlap the observed steps, which is why the correction holds across them.
What to do
For the number of available steps, ten is usually enough to stop the point estimate wandering, and somewhere between twenty and a hundred stabilises the standard error against a target you can check by redrawing. Past that you are spending computer time for nothing. For the distribution, either fit the tentative kernel to the observed steps and accept a basic model’s dependence on that choice, or carry movement covariates and let the integrated model take the choice off the table. Whichever you pick, the fitted model still needs checking against the data it did not see, which is the final post in this thread.
References
Northrup JM, Hooten MB, Anderson CR, Wittemyer G 2013. Ecology 94(7):1456-1463 (10.1890/12-1688.1).
Forester JD, Im HK, Rathouz PJ 2009. Ecology 90(12):3554-3565 (10.1890/08-0874.1).
Avgar T, Potts JR, Lewis MA, Boyce MS 2016. Methods in Ecology and Evolution 7(5):619-630 (10.1111/2041-210X.12528).
Thurfjell H, Ciuti S, Boyce MS 2014. Movement Ecology 2:4 (10.1186/s40462-014-0004-7).
Fortin D, Beyer HL, Boyce MS, Smith DW, Duchesne T, Mao JS 2005. Ecology 86(5):1320-1330 (10.1890/04-0953).
Fieberg J, Signer J, Smith B, Avgar T 2021. Journal of Animal Ecology 90(5):1027-1043 (10.1111/1365-2656.13441).