set.seed(132)
Tyr <- 25; phi_true <- 0.55; f_true <- 0.48; sy_true <- 11; N1_true <- 180
N <- numeric(Tyr); N[1] <- N1_true
for (t in 1:(Tyr - 1)) {
N[t + 1] <- rbinom(1, N[t], phi_true) + rpois(1, f_true * N[t])
}
C <- round(rnorm(Tyr, N, sy_true))
p_true <- 0.5; Kcr <- 9; Rrel <- rep(30, Kcr - 1)
recaps <- vector("list", Kcr - 1); never <- integer(Kcr - 1)
for (i in 1:(Kcr - 1)) {
alive <- rep(TRUE, Rrel[i]); fr <- integer(Rrel[i]); rc <- numeric(Kcr)
for (t in (i + 1):Kcr) {
alive <- alive & (runif(length(alive)) < phi_true)
got <- alive & (runif(length(alive)) < p_true) & (fr == 0); fr[got] <- t
}
for (t in (i + 1):Kcr) rc[t] <- sum(fr == t)
recaps[[i]] <- rc; never[i] <- Rrel[i] - sum(fr > 0)
}
Jyr <- 10; prodB <- rep(40, Jyr); prodJ <- rpois(Jyr, prodB * f_true)
kf_ll <- function(phi, f, sy, N1, C) {
lam <- phi + f; dv <- phi * (1 - phi) + f; a <- N1; P <- (2 * sy)^2; ll <- 0
for (t in seq_along(C)) {
if (t > 1) { ap <- a; a <- lam * ap; P <- lam^2 * P + dv * max(ap, 1) }
F <- P + sy^2; ll <- ll + dnorm(C[t], a, sqrt(F), log = TRUE)
K <- P / F; a <- a + K * (C[t] - a); P <- (1 - K) * P
}
ll
}
marray_ll <- function(phi, p) {
ll <- 0
for (i in 1:(Kcr - 1)) {
ts <- (i + 1):Kcr; q <- phi^(ts - i) * (1 - p)^(ts - i - 1) * p
ll <- ll + sum(recaps[[i]][ts] * log(q)) + never[i] * log(1 - sum(q))
}
ll
}Implicit information in integrated models
R
population dynamics
capture-recapture
demography
ecology tutorial
How an integrated model estimates a demographic rate it has no direct data for, how much each data set contributes, and why implicit estimates lean on it.
The previous post built an integrated population model (IPM) and showed that counts alone cannot separate adult survival from recruitment: the two lie anywhere along a ridge of constant growth rate. Adding capture-recapture and productivity data cut across that ridge and fixed both rates. This post looks at the mechanism more closely, and at a result that seems to give something for nothing: an IPM can estimate a rate for which it holds no direct data at all.
The reason is the shared trajectory. The counts fix the growth rate, which is survival plus recruitment. So if any one of the two rates is known, the other follows by subtraction. Capture-recapture data fix survival directly, and the counts then imply recruitment; a productivity survey fixes recruitment directly, and the counts then imply survival. This implicit information is real and useful, but it rests entirely on the demographic model being correct, and that is where the care comes in.
Estimating a rate with no data for it
We simulate the same kind of population as before: annual counts, a capture-recapture study on adults, and a productivity survey. Then we fit the IPM four ways, dropping data sets to see what each one identifies.
fitv <- function(use) {
has_p <- "cr" %in% use
init <- if (has_p) c(qlogis(.5), log(.5), log(12), qlogis(.5), log(C[1]))
else c(qlogis(.5), log(.5), log(12), log(C[1]))
fn <- function(par) {
phi <- plogis(par[1]); ff <- exp(par[2]); sy <- exp(par[3])
if (has_p) { p <- plogis(par[4]); N1 <- exp(par[5]) } else { N1 <- exp(par[4]) }
ll <- 0
if ("count" %in% use) ll <- ll + kf_ll(phi, ff, sy, N1, C)
if ("cr" %in% use) ll <- ll + marray_ll(phi, p)
if ("prod" %in% use) ll <- ll + sum(dpois(prodJ, prodB * ff, log = TRUE))
-ll
}
o <- optim(init, fn, method = "BFGS", hessian = TRUE,
control = list(maxit = 1500, reltol = 1e-11))
V <- solve(o$hessian); phi <- plogis(o$par[1]); ff <- exp(o$par[2])
c(phi = phi, f = ff, se_phi = sqrt(V[1, 1]) * phi * (1 - phi), se_f = sqrt(V[2, 2]) * ff)
}
cr <- fitv(c("count", "cr")) # counts + capture-recapture (no productivity)
prod <- fitv(c("count", "prod")) # counts + productivity (no capture-recapture)
full <- fitv(c("count", "cr", "prod"))Fit the counts together with the capture-recapture data but no productivity survey. Recruitment f is estimated at 0.524 (SE 0.045, true 0.48), even though not one recruit was counted: the capture-recapture data give survival, the counts give the growth rate, and recruitment falls out as the difference. The mirror image holds when we fit the counts with the productivity survey but no capture-recapture data: adult survival comes out at 0.537 (SE 0.037, true 0.55), with no marked animal ever re-encountered.
How much each data set contributes
The implicit estimates are not free of cost; they carry more uncertainty than a rate that has a data set speaking to it directly, and the full model, using every source, is the most precise of all.
sedf <- data.frame(
model = rep(c("count + CR", "count + productivity", "full"), each = 2),
rate = rep(c("survival", "recruitment"), 3),
se = c(cr["se_phi"], cr["se_f"], prod["se_phi"], prod["se_f"], full["se_phi"], full["se_f"])
)
sedf$model <- factor(sedf$model, levels = c("count + CR", "count + productivity", "full"))
ggplot(sedf, aes(rate, se, colour = model, group = model)) +
geom_line(position = position_dodge(0.35), linewidth = 0.5, alpha = 0.6) +
geom_point(position = position_dodge(0.35), size = 3.4) +
scale_colour_manual(values = c("count + CR" = sage,
"count + productivity" = gold, "full" = forest)) +
labs(x = NULL, y = "standard error", colour = NULL,
title = "What each data set buys in precision",
subtitle = "the full model is tightest for both rates") +
theme_te() + theme(legend.position = "top")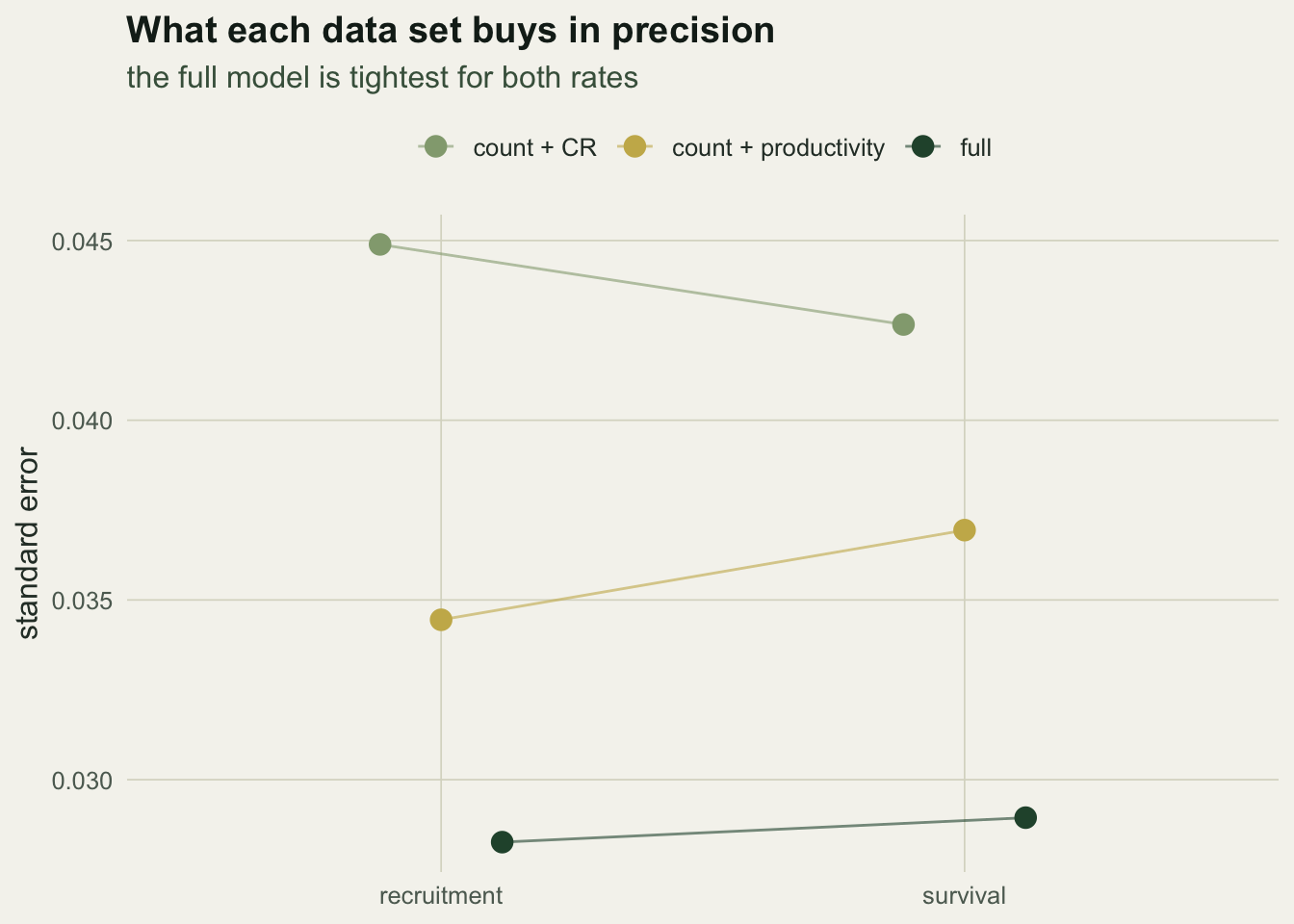
Here survival is estimated to SE 0.037 implicitly from counts and productivity, close to the SE 0.043 the capture-recapture data give directly; the full model tightens it to 0.029. The geometry behind this is the ridge from the previous post. Each data set adds a constraint that cuts across it, and where the constraints meet is where the rates are pinned.
syf <- exp(log(11)); N1f <- C[1] # fixed nuisances for the surfaces
# refit full with a fixed-p variant to get p for the CR surface
oful <- optim(c(qlogis(.5), log(.5), log(12), qlogis(.5), log(C[1])), function(par) {
-(kf_ll(plogis(par[1]), exp(par[2]), exp(par[3]), exp(par[5]), C) +
marray_ll(plogis(par[1]), plogis(par[4])) +
sum(dpois(prodJ, prodB * exp(par[2]), log = TRUE)))
}, method = "BFGS", control = list(maxit = 1500, reltol = 1e-11))
syf <- exp(oful$par[3]); N1f <- exp(oful$par[5]); pf <- plogis(oful$par[4])
pg <- seq(0.38, 0.72, length.out = 70); fg <- seq(0.30, 0.64, length.out = 70)
gr <- expand.grid(phi = pg, f = fg)
gr$count <- mapply(function(a, b) kf_ll(a, b, syf, N1f, C), gr$phi, gr$f)
gr$cr <- gr$count + sapply(gr$phi, function(a) marray_ll(a, pf))
gr$prod <- gr$count + sapply(gr$f, function(b) sum(dpois(prodJ, prodB * b, log = TRUE)))
gr$fulls <- gr$cr + sapply(gr$f, function(b) sum(dpois(prodJ, prodB * b, log = TRUE))) - gr$count
gr$rz <- gr$count - max(gr$count); gr$rz[gr$rz < -25] <- -25
lev <- function(z) max(z) - 2
ggplot(gr, aes(phi, f)) +
geom_raster(aes(fill = rz), interpolate = TRUE) +
scale_fill_gradient(low = paper, high = sage, name = "count log-lik") +
geom_contour(aes(z = cr), breaks = lev(gr$cr), colour = forest, linewidth = 0.7) +
geom_contour(aes(z = prod), breaks = lev(gr$prod), colour = gold, linewidth = 0.7) +
geom_contour(aes(z = fulls), breaks = lev(gr$fulls), colour = rust, linewidth = 0.7) +
geom_point(aes(x = phi_true, y = f_true), colour = rust, size = 3, shape = 17) +
annotate("text", x = 0.44, y = 0.62, label = "+ CR", colour = forest, size = 3.4) +
annotate("text", x = 0.68, y = 0.40, label = "+ productivity", colour = gold, size = 3.4) +
coord_equal() +
labs(x = "adult survival (phi)", y = "recruitment (f)",
title = "Each data set cuts the ridge",
subtitle = "capture-recapture pins survival, productivity pins recruitment") +
theme_te()Warning in geom_point(aes(x = phi_true, y = f_true), colour = rust, size = 3, : All aesthetics have length 1, but the data has 4900 rows.
ℹ Please consider using `annotate()` or provide this layer with data containing
a single row.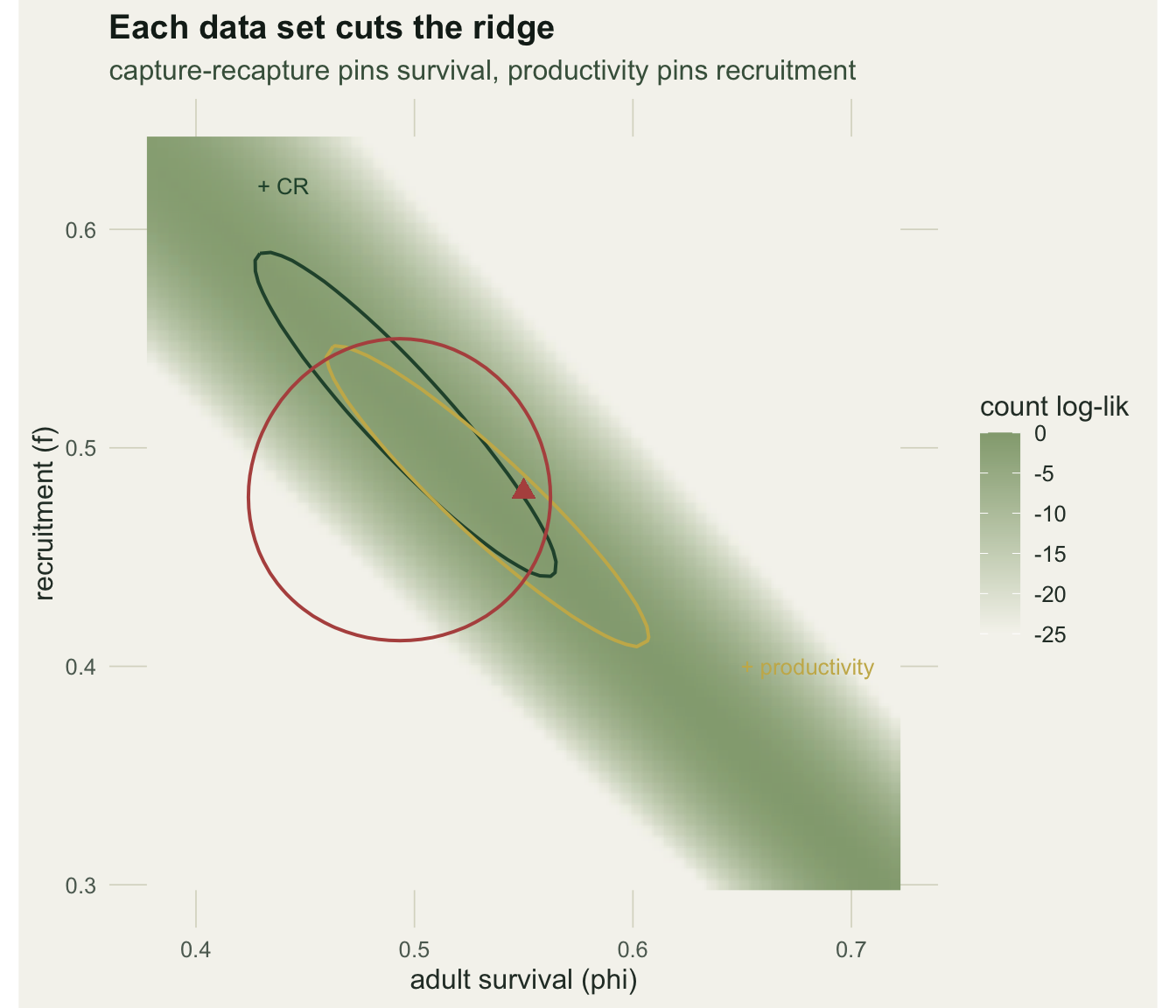
The catch: implicit means model-dependent
Implicit information comes from forcing the counts and the auxiliary data to agree through the demographic model. If that model is wrong, the implicit estimate absorbs the error. Suppose the productivity survey is biased high, as happens when the monitored nests are easier to find and more successful than average. Fit counts and the inflated productivity together, with no capture-recapture data to anchor survival.
prodJ_bias <- rpois(Jyr, prodB * f_true * 1.3) # productivity over-recorded by 30%
ob <- optim(c(qlogis(.5), log(.5), log(12), log(C[1])), function(par) {
-(kf_ll(plogis(par[1]), exp(par[2]), exp(par[3]), exp(par[4]), C) +
sum(dpois(prodJ_bias, prodB * exp(par[2]), log = TRUE)))
}, method = "BFGS", control = list(maxit = 1500, reltol = 1e-11))
phi_bias <- plogis(ob$par[1]); f_bias <- exp(ob$par[2])The inflated productivity pushes recruitment up to 0.61, and because the counts fix the growth rate, the implicit survival is dragged down to 0.402, well below the true 0.55. Nothing in this fit looks wrong: the counts are matched and recruitment agrees with its (biased) survey. The error only shows if a second source disagrees. Here the capture-recapture data would put survival near 0.513, so the implicit and the direct survival estimates conflict, and that conflict is the signal that a demographic assumption has failed. Diagnosing this kind of hidden disagreement between data sets is the subject of the checking post later in this series.
Takeaways
An integrated population model shares the growth rate across its data sets, so knowing one vital rate implies the other. That is why the model can report a survival probability with no marked animals and a recruitment rate with no offspring counts: the missing rate is recovered implicitly from the trajectory. The implicit estimate is genuine, and its precision can rival a direct estimate from a modest auxiliary survey, but it is only as trustworthy as the process model it leans on. A biased productivity survey quietly biases the implicit survival, and the giveaway is a clash with a second, direct estimate. The next post turns to a different question the IPM answers well: separating the year-to-year variability in the rates from the demographic noise of a finite population.
References
Besbeas, P., Freeman, S. N., Morgan, B. J. T. and Catchpole, E. A. 2002. Integrating mark-recapture-recovery and census data to estimate animal abundance and demographic parameters. Biometrics 58(3):540-547 (10.1111/j.0006-341X.2002.00540.x).
Schaub, M. and Abadi, F. 2011. Integrated population models: a novel analysis framework for deeper insights into population dynamics. Journal of Ornithology 152(S1):227-237 (10.1007/s10336-010-0632-7).
Abadi, F., Gimenez, O., Arlettaz, R. and Schaub, M. 2010. An assessment of integrated population models: bias, accuracy and violation of the assumption of independence. Ecology 91(1):7-14 (10.1890/08-2235.1).
Riecke, T. V., Williams, P. J., Behnke, T. L., Gibson, D., Leach, A. G., Sedinger, B. S., Street, P. A. and Sedinger, J. S. 2019. Integrated population models: model assumptions and inference. Methods in Ecology and Evolution 10(7):1072-1082 (10.1111/2041-210X.13195).
Zipkin, E. F. and Saunders, S. P. 2018. Synthesizing multiple data types for biological conservation using integrated population models. Biological Conservation 217:240-250 (10.1016/j.biocon.2017.10.017).
Kery, M. and Schaub, M. 2012. Bayesian Population Analysis using WinBUGS. Academic Press (ISBN 978-0-12-387020-9).